在自噬研究领域,ATG蛋白介导的泛素样偶联系统与吞噬泡的成核、延伸机制已得到详尽阐释。然而,一个长期存在的核心谜题在于:吞噬泡如何完成其末端的物理性切割与密封,从而形成一个完整的双膜自噬体?传统模型曾推测ATG蛋白本身或通过招募其他膜动态调控蛋白足以完成此步骤,而近年来针对这一谜题的研究明确指向一个进化保守、功能更为广泛的细胞机器——ESCRT-III复合物。继上一期对自噬“明星”家族ATGs的详尽介绍后,本期将聚焦于ESCRT-III复合物这一自噬体闭合的关键参与者,系统性介绍其在自噬中的定位与功能,并逐一解析ESCRT-III各成员的特征及其在疾病发生发展中的潜在意义。

一、ESCRT与ESCRT-III的细胞生物学功能
细胞内膜系统的动态重塑是维持细胞稳态的关键过程,而转运必需内体分选复合物(Endosomal sorting complex required for transport, ESCRT)是这个过程的核心角色之一。ESCRT是由多个亚基复合体(ESCRT-0、-I、-II、-III)及辅助蛋白(如Vps4、Vta1等)组成的精密分子机器,其核心功能是介导细胞膜与膜结合细胞器的“由内向外”膜重塑与切割事件。ESCRT通路最初在酵母中发现,因其参与将泛素化膜蛋白分选至内吞体腔内的囊泡(即多泡体形成)而得名[1],但后续研究揭示其功能远不止于此,ESCRT复合体,尤其是其终端执行模块ESCRT-III,以其独特的逆拓扑膜剪接能力,广泛参与多种细胞进程,如细胞分裂、细胞因子脱落、病毒出芽、细胞器修复及自噬等多个关键的细胞生物学过程[2]。
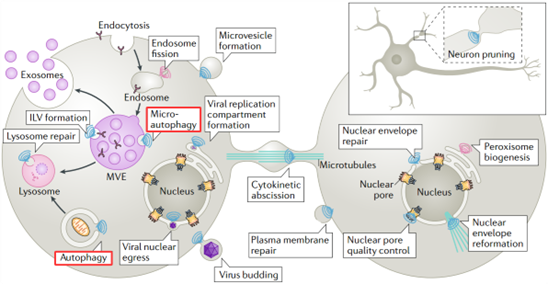
图1. ESCRT介导的生物学过程概述[2]
作为ESCRT系统中的最终执行者,ESCRT-Ⅲ由带正电的细胞质蛋白(在人类中主要为CHMP家族蛋白)构成,这些蛋白通常以非活跃单体形式存在,在被ESCRT-I/II或其他适配蛋白(如ALIX)招募至特定膜位点后,发生构象改变并聚合形成带状螺旋聚合物[3]。这些聚合物在膜内侧组装成紧缩环或螺旋结构,通过消耗ATP(间接由Vps4提供能量)发生构象变化,产生使膜颈部收缩和切割的机械力[2]。因此,ESCRT-III的核心细胞生物学功能是直接驱动膜变形、膜颈收缩和最终的膜切割反应,这是多种膜脱离过程共同的终末步骤。这一特性使其成为解决自噬体“开合问题”的理想候选者。现已有大量文献证实,在经典自噬及多种选择性自噬(如线粒体自噬、内质网自噬)过程中,ESCRT-III的核心组分(如CHMP4B、CHMP2A)及其AAA+ATP酶调节因子VPS4,被特异性招募至即将闭合的自噬体颈部,并施加机械力完成膜分离[4]。
二、ESCRT-III的主要成员解析
ESCRT-III并非一个静态复合物,而是由多个带正电的、小分子量带电多囊体蛋白(Charged Multivesicular Body Protein, CHMP)以及辅助蛋白(如IST1)动态组装而成的多聚体。这些成员在蛋白结构上共享以下高度保守的经典架构,这决定了其独特的工作模式—从自抑制的胞质单体,转变为在膜上活跃组装的“分子机器”。其核心结构域主要包括:
1. N端碱性结构域:通常由带正电荷的氨基酸组成,是蛋白与带负电荷的膜磷脂(如磷脂酰丝氨酸)结合的关键区域,负责将ESCRT-III锚定至目标膜位点。
2. C端酸性结构域:富含带负电荷的氨基酸。在胞质单体状态下,该结构域会反向与N端碱性结构域相互作用,形成一种“闭合”的自抑制构象,阻止蛋白发生非特异性聚集和膜结合,确保其仅在被正确招募后才被激活。
3. MIT相互作用基序(MIM):多数ESCRT-III成员(如CHMP1-3、IST1)在其序列中包含一个或多个短线性基序,即MIM。该基序可被AAA+ATP酶VPS4的MIT结构域特异性识别,是VPS4被招募至ESCRT-III多聚体并驱动其拆卸和重塑的结构基础。
ESCRT-III成员的自抑制状态可被上游信号(如ESCRT-II或ALIX)解除。激活后,蛋白发生构象变化,暴露出多聚化界面,通过“头对尾”或侧向相互作用,组装成螺旋细丝或多聚环,直接推动膜变形。总之,ESCRT-III成员通过其保守的“自抑制单体→膜结合→构象开放→多聚组装”的结构逻辑,形成了一个可被精准调控的通用膜重塑模块[2, 5-6]。
ESCRT-III的主要成员情况如下:
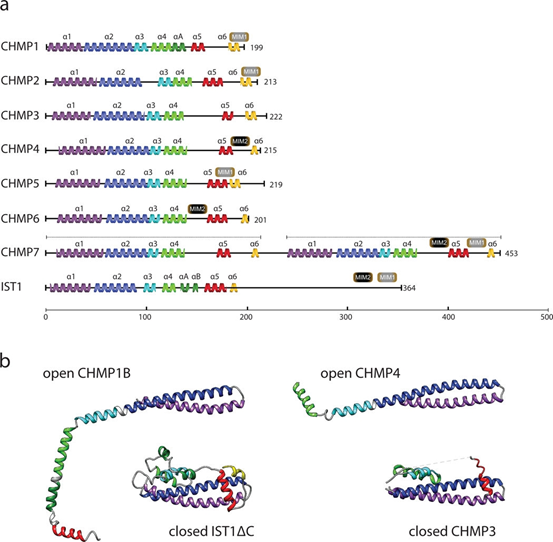
图2. ESCRT-III蛋白结构示意图[5]
CHMP4:ESCRT-III中最主要和含量最丰富的成丝蛋白,包括CHMP4A、B、C等多个旁系同源物。CHMP4构成多聚体的核心,直接参与膜结合和变形。CHMP4C在细胞分裂退出调控中扮演特殊角色,其磷酸化状态受Aurora B激酶调控,从而参与细胞质分裂检查点,防止染色体分离错误时发生异常脱落。
CHMP3与CHMP2:这两种蛋白通常形成异源二聚体,并与CHMP4细丝侧向结合,被认为在调节细丝几何形状、稳定多聚体结构以及招募VPS4方面起关键作用。它们与VPS4的MIT结构域相互作用,促进ESCRT-III亚基的周转和复合体的最终解离。
CHMP1(CHMP1A/B)与IST1:这一对蛋白可以形成独特的双链螺旋共聚物,能够结合并重塑正弯曲膜(与经典的逆拓扑膜作用相反)。此外,IST1也存在于ESCRT-III的核心组装中,可能起到调节复合体功能或提供额外连接位点的作用。在自噬相关背景下,它们通过IST1招募微管切割ATP酶spastin,协调自噬体成熟或细胞器周转中的微管动力学。
CHMP6:通常作为ESCRT-III的成核因子,被ESCRT-II的EAP20亚基招募,进而激活CHMP4的聚合。它在经典的多泡内体通路中作用明确,但在自噬体闭合中的必要性可能因细胞环境和招募路径的不同而存在差异。
CHMP7:一种特殊的ESCRT-III相关蛋白,N端具有类似ESCRT-II的翼状螺旋结构域,C端则类似CHMP6。它主要参与核膜相关事件,如有丝分裂后核膜封闭和核孔复合体质量监控。虽然不直接参与经典自噬,但其功能体现了ESCRT-III在膜密封中的普遍原理。CHMP7通过内核膜蛋白LEMD2被招募至核膜,进而募集CHMP4B等核心ESCRT-III组分。
CHMP5:亦称Vps60,作为辅助蛋白在功能上更倾向于调节者角色。CHMP5通过其富含的MIT作用基序高效招募并激活VPS4 ATP酶,是连接ESCRT-III多聚体与其拆卸引擎的关键适配器,对调控整个机器的周转效率至关重要。然而,目前证据显示其在自噬体闭合中并非直接执行者,其影响可能更多通过调节VPS4活性间接实现。
研究表明,ESCRT-III成员的功能障碍与多种人类疾病密切相关。CHMP1A的双等位基因变异则会导致脑桥小脑发育不全8型;CHMP2B的显性突变已被发现是额颞叶痴呆和和肌萎缩侧索硬化的病因之一,这提示内体—溶酶体通路及膜运输障碍在神经退行性疾病中的重要性;CHMP3纯合子变异与遗传性痉挛性截瘫相关;CHMP4C功能异常与基因组不稳定性及癌症风险相关;此外,ESCRT-III在病毒(如HIV、疱疹病毒)出芽中的关键作用,使其成为潜在的抗病毒治疗靶点。而在自噬方面,虽然尚未有ESCRT-III核心成员突变直接导致纯自噬缺陷病的广泛报道,但考虑到其在自噬体闭合中的必需功能,其在神经退行性疾病(如阿尔茨海默病、帕金森病)以及癌症等与自噬紊乱密切相关的疾病病理过程中可能扮演着调节者或协同因素的角色[7]。
三、ESCRT-III在自噬体闭合中的核心作用
自噬作为一种高度保守的降解途径,通过形成双层膜结构的自噬体包裹待降解物质,并将其递送至溶酶体。自噬体的形成始于吞噬泡,在内质网等相关膜结构附近形成并延伸,逐渐弯曲成杯状结构,包裹待降解的胞质内容物。随着吞噬泡边缘的不断延伸并逐渐靠拢,其开口逐渐缩小,最终需要通过膜剪接事件实现完全密封,从而形成一个独立、闭合的双层膜自噬体。自噬体的闭合是自噬通路中一个关键的限速与质检步骤,可以确保待降解的胞质成分被完全密封于双层膜的自噬体内,从而为后续与溶酶体的特异性融合奠定基础。在哺乳动物细胞中,这一精密的膜剪接事件主要由ESCRT-III蛋白复合体及其协同的AAA-ATP酶VPS4所驱动[2]。
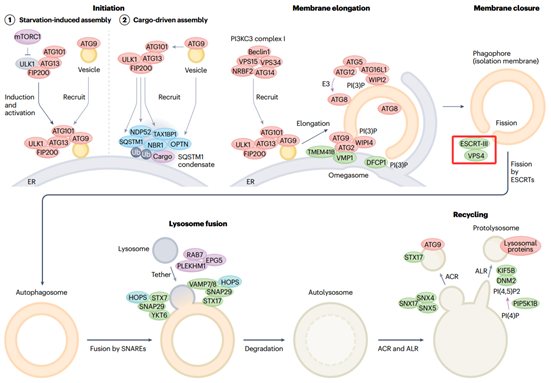
图3. 大自噬的膜动力学[4]
在自噬过程中,核心的ESCRT-III组分(如CHMP4B、CHMP2A、CHMP3等)及VPS4被特异地、瞬时地募集至即将闭合的吞噬泡边缘[7]。被招募后,ESCRT-III蛋白在膜颈部的胞质面发生构象转变,从自抑制的单体聚合成螺旋状的细丝或多聚体环。这些结构作为物理性的“收缩装置”,紧密结合在膜下方。随后,VPS4被募集至ESCRT-III多聚体上,通过水解ATP提供能量,驱动ESCRT-III亚基的动态周转与多聚体重塑,从而对膜颈产生持续的机械力,使其逐步收缩并最终完成膜剪接[2, 4, 8]。ESCRT-III功能缺失时,吞噬泡将无法完成最终密封,形成有结构性缺陷的“未闭合自噬体”。这些异常结构无法正常招募介导自噬体-溶酶体融合的关键SNARE蛋白(如STX17),最终导致自噬流整体中断,并可能累积异常的膜结构,影响细胞稳态。因此,ESCRT-III介导的闭合是自噬体获得功能成熟、进入下游降解程序的必经之门。
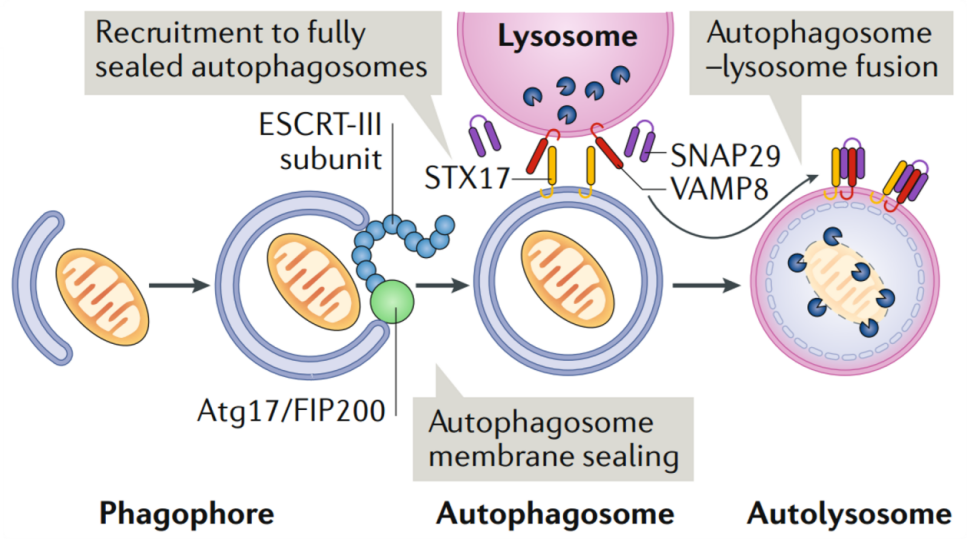
图4. 大自噬过程中ESCRT介导的吞噬细胞的封闭[2]
综上所述,ESCRT-III是一个多功能、动态变化的膜重塑机器,其通过介导逆拓扑膜剪接,在包括自噬在内的众多基础细胞生物学过程中发挥核心作用。在自噬中,ESCRT-III是确保自噬体正确闭合,从而维持自噬流畅通的关键执行者。从神经退行性疾病、癌症到感染性疾病,ESCRT-III的功能异常或病毒对其的劫持都可能成为疾病发生发展的环节。对ESCRT-III功能的深入研究,不仅加深了我们对自噬分子机制的理解,也揭示了其在人类健康与疾病中的广泛意义。
本期我们深入解析了ESCRT-III 复合物如何作为自噬体闭合的“终极剪接师”,确保待降解内容物被完整封装。然而,闭合只是故事的开端,一个完美密封的自噬体,又该如何准确找到溶酶体并与之融合,释放内容物进行降解呢?这就需要另一组关键蛋白的引导:SNARE复合物。下一期,我们将聚焦介导自噬体—溶酶体融合的核心“对接密码”——STX17及其家族基因,继续拆解自噬这条精密供应链,敬请关注。
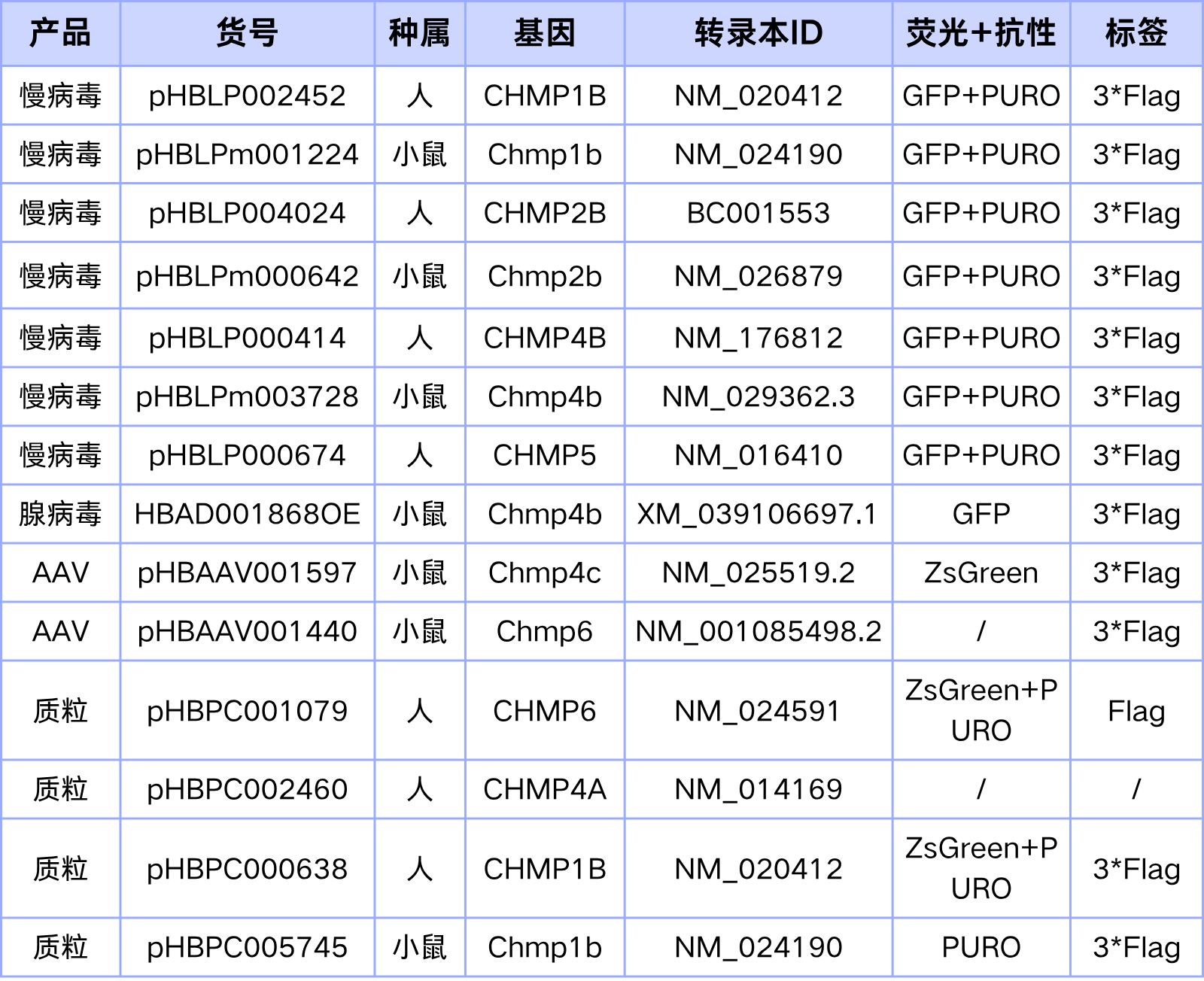
汉恒生物专营病毒包装十余载,建立了庞大的基因研究现货工具库,现可提供ESCRT-III相关基因的表达调控工具(见表1),也可定制靶向特异性组织或细胞的基因调控工具,包括慢病毒(Lentivirus, LV)、腺病毒(Adenovirus, AD)、腺相关病毒(Adeno-associated-virus, AAV)以及质粒等,如有技术或产品需求,欢迎随时咨询汉恒生物微信公众号或拨打官网技术服务热线:400-092-0065。
表1. 汉恒生物ESCRT-III 相关基因调控的现货
参考文献:
[1] Katzmann DJ, Babst M, Emr SD. Ubiquitin-dependent sorting into the multivesicular body pathway requires the function of a conserved endosomal protein sorting complex, ESCRT-I. Cell. 2001 Jul 27;106(2):145-55.
[2] Vietri M, Radulovic M, Stenmark H. The many functions of ESCRTs. Nat Rev Mol Cell Biol. 2020 Jan;21(1):25-42.
[3] Bajorek M, Schubert HL, McCullough J, Langelier C, Eckert DM, Stubblefield WM, Uter NT, Myszka DG, Hill CP, Sundquist WI. Structural basis for ESCRT-III protein autoinhibition. Nat Struct Mol Biol. 2009 Jul;16(7):754-62.
[4] Yamamoto H, Zhang S, Mizushima N. Autophagy genes in biology and disease. Nat Rev Genet. 2023 Jun;24(6):382-400.
[5] Schöneberg J, Lee IH, Iwasa JH, Hurley JH. Reverse-topology membrane scission by the ESCRT proteins. Nat Rev Mol Cell Biol. 2017 Jan;18(1):5-17.
[6] McCullough J, Clippinger AK, Talledge N, Skowyra ML, Saunders MG, Naismith TV, Colf LA, Afonine P, Arthur C, Sundquist WI, Hanson PI, Frost A. Structure and membrane remodeling activity of ESCRT-III helical polymers. Science. 2015 Dec 18;350(6267):1548-51.
[7] Hurley JH, Coyne AN, Miączyńska M, Stenmark H. The expanding repertoire of ESCRT functions in cell biology and disease. Nature. 2025 Jun;642(8069):877-888.
[8] Zhou F, Wu Z, Zhao M, Murtazina R, Cai J, Zhang A, Li R, Sun D, Li W, Zhao L, Li Q, Zhu J, Cong X, Zhou Y, Xie Z, Gyurkovska V, Li L, Huang X, Xue Y, Chen L, Xu H, Xu H, Liang Y, Segev N. Rab5-dependent autophagosome closure by ESCRT. J Cell Biol. 2019 Jun 3;218(6):1908-1927.









 8434
8434










