前几期文章我们已系统介绍了ULK、WIPI、ATG、CHMP等基因家族在自噬过程中的关键作用。其中,ULK通过形成ULK复合物与mTOR受体相互作用启动自噬;WIPI和ATG均为调控自噬小体形成的关键基因;CHMP则参与自噬小体的闭合过程。那么,自噬小体在闭合完成后,是通过怎样的分子机制与溶酶体融合,最终形成自噬溶酶体,从而完成整个自噬过程的呢?本期我们将聚焦于促进自噬溶酶体形成的关键基因——syntaxins(STX)家族,深入探讨其在自噬溶酶体融合步骤中的核心作用。

STX与SNARE的生物学功能
SNARE是一类关键的膜融合因子,对细胞内多种膜融合途径至关重要。STX作为SNARE蛋白家族的一个重要亚类,在人类基因组中有15个成员(STX1、STX2、STX3、STX4、STX5、STX6、STX7、STX8、STX10、STX11、STX12、STX16、STX17、STX18、STX19)。它们通过其保守的SNARE结构域与其他SNARE蛋白(如SNAP29、VAMP等)组装成SNARE复合物,拉近并催化两个脂质双层的融合。
早期研究曾根据定位差异将SNARE蛋白分为v-SNARE(定位于囊泡膜)或t-SNARE(定位于靶膜),STX基因家族最初被归类为t-SNARE[1]。然而,随着研究的深入,这种严格的二元分类被证明无法涵盖所有的膜融合事件,某些SNARE蛋白在不同膜融合过程中与不同蛋白相互作用。
因此,研究人员提出以SNARE蛋白核心结构域的构象特征和膜融合过程中自由能的变化为依据,将其分为为Qa-、Qb-、Qc-和R-SNARE四类[2]。Qa-、Qb-和Qc-SNARE含有单个SNARE基序,基序中心为保守的谷氨酰胺(Q)残基,Qa-SNARE以及部分Qb-、Qc-SNARE的N端含有三个反向平行的螺旋束;R-SNARE含有单个SNARE基序,基序中心为保守的精氨酸(R)残基。STX基因家族成员中,STX6、STX8、STX10为Qc-SNARE,其余均为Qa-SNARE。
驱动膜融合的功能性SNARE复合物通常由这四个类型各一个成员通过一个灵活的连接体与SNARE基序相连,组成具有平行四螺旋结构的异源寡聚体[3]。有一类特殊SNARE蛋白SNAP,同时拥有Qb-和Qc-SNARE基序,归类于Qbc-SNARE,这类SNARE与一个Qa-SNARE和一个R-SNARE结合即可组成SNARE复合体。
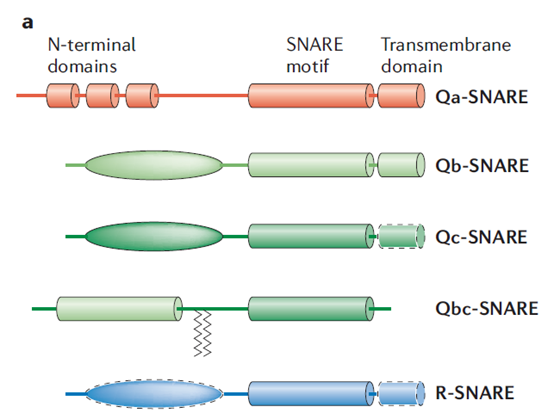
图1. 不同SNARE蛋白的结构[4]
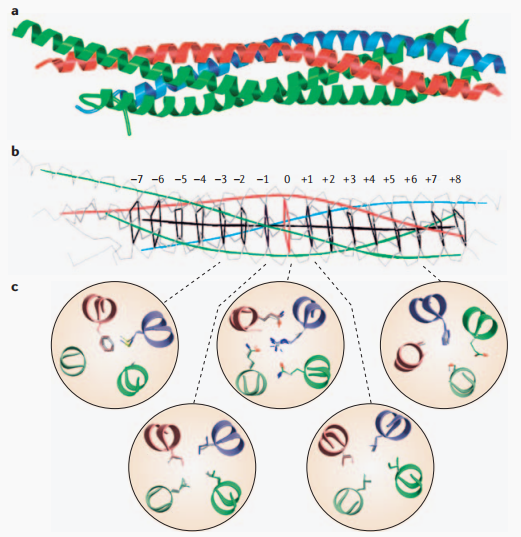
图2. SNARE复合物结构示意图[4]
目前,多个STX家族成员在哺乳动物细胞中的功能已得到证实,它们均参与SNARE复合物的形成,促进囊泡的融合:STX7-VTI1b-STX8-VAMP7/ VAMP8在内体的成熟过程中促进囊泡的融合,最终形成功能性的溶酶体;STX18-SEC20-USE1/SLT1-SEC22b参与内质网与高尔基体之间的囊泡运输;STX5-GS27-BET1-SEC22b可将囊泡从内质网运输至高尔基体;STX13(也认为是STX12)/STX16-VTI1a-STX6-VAMP4参与早期分选内体的形成;STX2/STX4-SNAP23-VAMP2/synaptobrevin 2/ cellubrevin/VAMP7参与再循环内体的形成;STX1-SNAP25-VAMP2/ synaptobrevin 2和STX2-SNAP23-VAMP8两种SNARE复合物均参与细胞内囊泡运输和细胞膜融合过程,确保神经递质能够从囊泡中释放到突触间隙,实现神经信号的传递(图3)[4]。
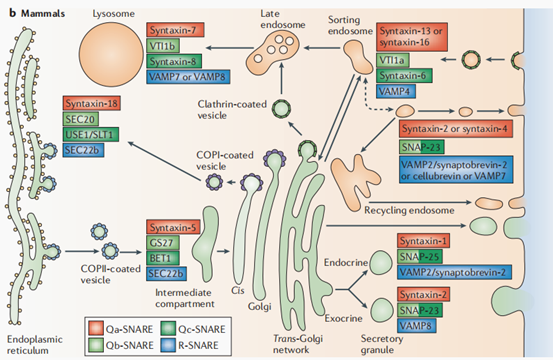
图3. 哺乳动物细胞中不同SNARE复合物在细胞内膜融合中的作用[4]
与自噬相关的STX基因
STX基因家族在细胞内囊泡运输和膜融合过程中发挥至关重要的作用,然而目前已知参与自噬过程的成员较少。研究表明,STX17和STX7参与自噬小体-溶酶体融合过程,而STX13则参与自噬小体的形成过程,其他STX家族成员暂无自噬相关研究报导。接下来,本文将详细介绍STX17、STX7和STX13在自噬过程中的具体作用机制。
STX17:自噬小体-溶酶体融合的关键调控因子
自噬小体与溶酶体融合形成自噬溶酶体,是自噬降解过程的关键过程,溶酶体必须与完全闭合的自噬小体融合,被吞噬物质才会顺利降解,STX17是这一过程的重要介质。
从结构上看,STX17包含一个位于N端的Habc结构域、一个Qa-SNARE基序,以及C端附近的两个串联跨膜结构域。在非激活状态下,Habc结构域与Qa-SNARE基序相互作用使STX17以自抑制的闭合构象存在。值得注意的是,STX17的Qa-SNARE区域内含有一个LC3相互作用基序(LIR基序),ATG8家族成员GABARAP可通过与Habc结构域竞争结合Qa-SNARE基序,解除其自抑制状态,促使STX17转变为开放构象,从而启动SNARE复合物的组装[5]。C端两个跨膜结构域则可以通过甘氨酸-拉链基序形成发夹状结构,这一特征结构是STX17能够特异性靶向已完成闭合的自噬小体外膜的关键[6]。
在自噬小体闭合后,STX17会被招募到自噬小体膜上,随后STX17与溶酶体膜中的VAMP7或VAMP8(R-SNARE)以及SNAP29(Qbc-SNAREs)形成复合物,以促进自噬小体-溶酶体融合。融合完成后,随着自噬小体膜的降解,STX17从膜上解离[7]。
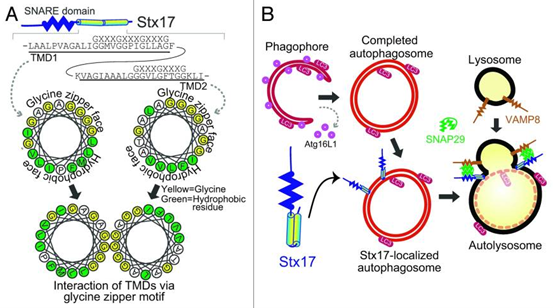
图4. STX17介导自噬小体-溶酶体融合[8]
STX17-SNAP29-VAMP8复合物的形成过程由ATG14调控,ATG14与STX17相互作用,稳定STX17和SNAP29形成的螺旋结构,促进SNARE复合物的形成。此外,E3泛素连接酶MARCH7能够介导ATG14泛素化,抑制其与STX17结合,影响SNARE复合物的形成,进而抑制自噬,这一调控通路的失调已被证实与神经退行性疾病等多种自噬相关疾病密切相关[9]。
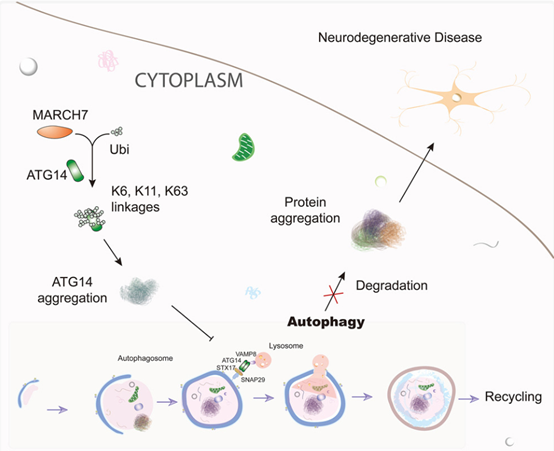
图5. ATG14泛素化抑制自噬[9]
STX7:独立于STX17的自噬融合调控者
如前所述,STX17-SNAP29-VAMP8复合物在自噬溶酶体形成中发挥关键作用。除此之外,STX7也有研究证明其参与该过程,Takahide Matsui等[10]发现YKT6可与SNAP29及溶酶体定位的STX7组装形成功能性SNARE复合物,有效驱动自噬小体与溶酶体的融合。值得注意的是,在STX17敲除细胞中,自噬小体-溶酶体融合及自噬溶酶体形成仍可稳定进行,表明STX7-SNAP29-YKT6复合物能够独立于经典的STX17-SNAP29-VAMP8途径发挥作用。这两套SNARE机制的存在,为细胞自噬流提供了重要的功能冗余。
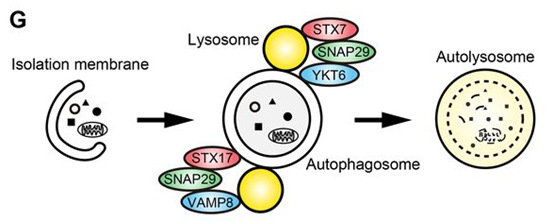
图6. 自噬小体与溶酶体融合过程示意图[10]
STX13:自噬小体成熟与分泌颗粒自噬的多面调控者
STX13于1988年首次被发现,它存在于吞噬体中,在调节所有细胞类型共有的基本膜转运过程方面起重要作用(图6)。
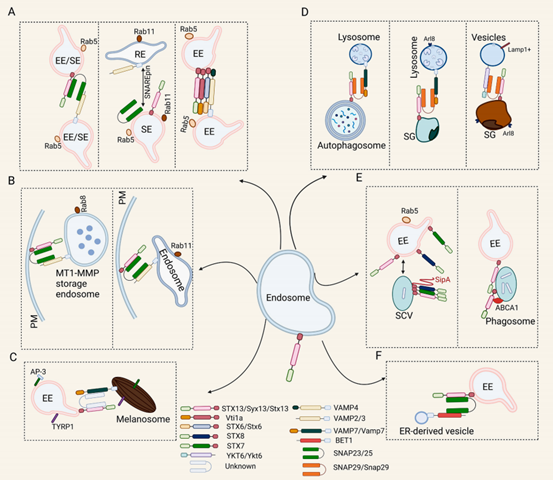
图7. STX13形成SNARE复合物介导膜融合。A:STX13-VAMP2-3-SNAP23/25和STX13-STX6-STX10-Vti1a复合物介导内体同型融合。B:STX13-VAMP2/3-SNAP23复合物介导内体-质膜融合。C:STX13-VAMP7-RSNARE复合物介导内体与黑素小体的融合。D:STX13-SNAP29-VAMP7或STX13-SNAP29- YKT6复合物介导分泌颗粒-自噬小体与溶酶体的融合。E:STX13影响吞噬体与内体融合;STX13-STX7-STX8-SipA复合体介导内体与含沙门氏菌液泡(scv)的融合。F:STX13-BET1-SNAP23复合物介导内质网源性囊泡与内体融合[11]。
在哺乳动物和果蝇细胞中,STX13与vti1a(v-SNARE)结合通过ESCRTIII影响细胞自噬小体的成熟[12]。此外,在果蝇中还存在一种特殊的自噬类型,即精细胞自噬,在这种情况下,分泌性颗粒(SGs)的多余部分通过与溶酶体直接融合而被降解,这是一种分泌颗粒特异性的自噬过程。在果蝇的唾液腺细胞中,STX13/SNAP29/VAMP7可能会形成SNARE复合物,该复合物通过促进Arl8+溶酶体与成熟SGs的融合来促进蛋白质降解[13]。在果蝇幼虫的唾液腺中,STX13/SNAP29/ YKT6复合物通过介导SGs与Lamp1+囊泡之间的持续融合,形成降解小体,有助于清除冗余的分泌物质[14]。综上,STX13通过组建不同的SNARE复合物,在经典自噬与组织特异性自噬过程中均扮演着不可或缺的角色。
综上所述,STX蛋白作为膜融合的“分子桥梁”,在维持细胞稳态中发挥着不可或缺的作用,其功能异常会引发神经退行性疾病、代谢性疾病等。因此,对STX家族基因的深入研究,不仅加深了我们对细胞内各类膜融合机制的理解,也为相关疾病的诊治提供了新的思路与靶点。
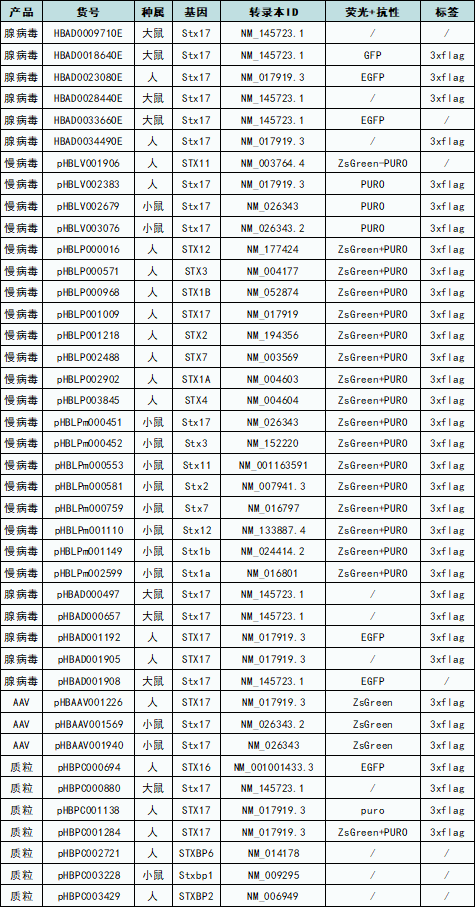
至此,自噬相关基因专题系列文章就结束了。汉恒生物专营病毒包装十余载,建立了庞大的基因研究现货工具库,现可提供STX相关基因的表达调控工具(见表1),也可定制靶向特异性组织或细胞的基因调控工具,包括慢病毒(Lentivirus, LV)、腺病毒(Adenovirus, AD)、腺相关病毒(Adeno-associated-virus, AAV)以及质粒等,如有技术或产品需求,欢迎随时咨询汉恒生物微信公众号或拨打官网技术服务热线:400-092-0065。
表1. 汉恒生物STX家族基因现货
参考文献:
[1]Söllner, T., Bennett, M. K., Whiteheart, S. W., Scheller, R. H., & Rothman, J. E. (1993). A protein assembly-disassembly pathway in vitro that may correspond to sequential steps of synaptic vesicle docking, activation, and fusion. Cell, 75(3), 409–418.
[2]Fasshauer, D., Sutton, R. B., Brunger, A. T., & Jahn, R. (1998). Conserved structural features of the synaptic fusion complex: SNARE proteins reclassified as Q- and R-SNAREs. Proceedings of the National Academy of Sciences of the United States of America, 95(26), 15781–15786.
[3]Fasshauer D. (2003). Structural insights into the SNARE mechanism. Biochimica et biophysica acta, 1641(2-3), 87–97.
[4]Jahn, R., & Scheller, R. H. (2006). SNAREs--engines for membrane fusion. Nature reviews. Molecular cell biology, 7(9), 631–643.
[5]Li, Y., Cheng, X., Li, M., Wang, Y., Fu, T., Zhou, Z., Wang, Y., Gong, X., Xu, X., Liu, J., & Pan, L. (2020). Decoding three distinct states of the Syntaxin17 SNARE motif in mediating autophagosome-lysosome fusion. Proceedings of the National Academy of Sciences of the United States of America, 117(35), 21391–21402.
[6]Itakura, E., Kishi-Itakura, C., & Mizushima, N. (2012). The hairpin-type tail-anchored SNARE syntaxin 17 targets to autophagosomes for fusion with endosomes/lysosomes. Cell, 151(6), 1256–1269.
[7]Tsuboyama, K., Koyama-Honda, I., Sakamaki, Y., Koike, M., Morishita, H., & Mizushima, N. (2016). The ATG conjugation systems are important for degradation of the inner autophagosomal membrane. Science (New York, N.Y.), 354(6315), 1036–1041.
[8]Itakura E, Mizushima N. Syntaxin 17: the autophagosomal SNARE. Autophagy. 2013 Jun 1;9(6):917-9.
[9]Shi, X., Wu, W., Feng, Z., Fan, P., Shi, R., & Zhang, X. (2023). MARCH7-mediated ubiquitination decreases the solubility of ATG14 to inhibit autophagy. Cell reports, 42(9), 113045.
[10]Matsui, T., Jiang, P., Nakano, S., Sakamaki, Y., Yamamoto, H., & Mizushima, N. (2018). Autophagosomal YKT6 is required for fusion with lysosomes independently of syntaxin 17. The Journal of cell biology, 217(8), 2633–2645.
[11]Zhang, W., Zhang, H., He, T., Li, X., Liu, Y., Han, H., & Wang, J. (2025). The diverse functions of syntaxin 13 in endosome-mediated membrane fusion. International journal of biological macromolecules, 307(Pt 1), 141878.
[12]Lu, Y., Zhang, Z., Sun, D., Sweeney, S. T., & Gao, F. B. (2013). Syntaxin 13, a genetic modifier of mutant CHMP2B in frontotemporal dementia, is required for autophagosome maturation. Molecular cell, 52(2), 264–271.
[13]Csizmadia, T., Lőrincz, P., Hegedűs, K., Széplaki, S., Lőw, P., & Juhász, G. (2018). Molecular mechanisms of developmentally programmed crinophagy in <i>Drosophila</i>. The Journal of cell biology, 217(1), 361–374.
[14]Szenci, G., Glatz, G., Takáts, S., & Juhász, G. (2024). The Ykt6-Snap29-Syx13 SNARE complex promotes crinophagy via secretory granule fusion with Lamp1 carrier vesicles. Scientific reports, 14(1), 3200.









 8434
8434










